Les écosystèmes sont de types variés mais leur nombre n'est pas si important que cela si l'on procède à quelques simplifications et à autant d'abstractions. Dans ce chapitre, je décrirai les formes générales de quatre d'entre eux, qui se laissent facilement observer : la forêt, le milieu aquatique terrestre (lac et étang, ou encore milieux lentiques), la rivière et la tourbière d'altitude. D'une part, seront à la fois rapprochées et comparées les forêts de nos contrées (forêt caducifoliée = à feuillage caduc, tempérée, biomes méditerranéens) et les forêts ombrophiles (à exigence forte en eau) tropicales ; d'autre part, il est difficile de séparer par leur fonctionnement respectif un lac et un étang, ce dernier étant le plus souvent un représentant du premier quand celui-ci arrive en de fin de vie : un lac vit et meurt, j'y reviendrai. Très différents sont les écosystèmes se rapportant à la rivière (milieu lotique), ou encore l'écosystème d'une tourbière de haute altitude, nous le verrons à la fin de ce chapitre. Dans le suivant, je décrirai le fonctionnement d'un écosystème qui se laisse beaucoup moins deviner que les précédents, à moins d'une observation fine et rapprochée, je vous parlerai du système vivant " sol " ainsi que de la vie et de la mort d'une simple feuille d'arbre...a
Toutes les forêts du monde ne se ressemblent pas, mais elles sont toutes représentatives d'écosystèmes supérieurement évolués, complexes et élaborés. Leur mécanisme de fonctionnement exige un minimum de précipitations annuelles (> 500 mm/an), ainsi que des températures supérieures à 10°C pendant 4 mois successifs. Bien évidemment, tout dépend de la latitude où l'on se trouve. Si l'on additionne l'ensemble des forêts de la planète (forêts boréales de conifères ou taïga, forêts caducifoliées tempérées, biomes méditerranéens, savanes et forêts tropicales ombrophiles), cela représente 28% de la surface du globe, soit 43 millions de km² ; à lui tout seul, cet ensemble produit 45% de la matière organique de la biosphère, soit les ¾ de la biomasse produite sur la terre ferme.
Chaque année, les forêts françaises produisent entre 10 et 20 tonnes de matière sèche par hectare, dont 1/3 sous forme de bois ; au total, les forêts de France accroissent leur volume de 60 millions de m³/an, et seulement une moitié est exploitée. La masse de carbone fixé par hectare oscille entre 3 et 4 t/an, soit un total de 70 millions de tonnes de carbone fixé par nos forêts chaque année, cependant que sont rejetés 140 millions de tonnes d'oxygène atmosphérique. Pour comparaison, une forêt tropicale fixe, elle, 10 à 20 t de carbone par hectare et par an, mais, compte tenu d'une respiration plus intense, elle subit une perte considérable, si bien qu'elle ne produit qu'une trentaine de tonnes de matière sèche par hectare et par an.

La croissance verticale a été conférée aux végétaux grâce à l'invention du bois (la lignine) ; on parle de végétaux ligneux. La forêt s'organise en strates fonctionnelles qui s'empilent les unes sur les autres ( stratification) ; les conditions biotiques et abiotiques varient de l'une à l'autre si bien que ce partage vertical permet tout à la fois de multiplier les niches écologiques (biodiversité), de limiter la concurrence (meilleur rendement), et d'exploiter du mieux possible l'énergie solaire incidente. On peut facilement imaginer la lutte sans merci auxquels s'adonnent les arbres et autres végétaux moins ligneux pour la lumière : la canopée, qui est la strate supérieure de la forêt, reçoit davantage d'énergie lumineuse que les strates sous-jacentes, les végétaux doivent donc s'adapter, vaincre ou dépérir.
Pyramide des biomasses d'une forêt de nos contrée
Physiologiquement, il faut considérer un arbre comme une pompe très puissante qui peut élever l'eau du sol de 1 à 7 m par heure dans son tronc. C'est surtout la force créée par l'évaporation foliaire qui permet une telle absorption racinaire. De même, l'arbre est une véritable usine à fabriquer de la matière organique (sucres principalement) ; l'eau du sol, chargée de sels minéraux (sève brute), monte des racines aux feuilles qui sont le siège de la photosynthèse : le gaz carbonique qu'elles absorbent est transformé en sucres grâce à l'énergie fournie par la lumière solaire. Ces sucres (glucides) repartent dans le flot de la circulation inverse (sève élaborée) et vont alimenter l'ensemble de l'arbre, tout en permettant à ce dernier de constituer ses réserves. Ainsi, chaque année, le diamètre d'un arbre s'accroît de 1 mm à 2 cm selon les conditions climatiques et la latitude.
Un écosystème forêt de feuillus tempérée représente une biomasse totale (producteurs + consommateurs + décomposeurs) allant de 300 à 400 t/ha. Comparée à une prairie constamment dévorée (60 à 80%) par les herbivores qu'elle produit, notre forêt ne contient qu'une dizaine de kilogrammes de consommateurs par hectare, ce qui représente une très faible consommation de végétaux dans le système (1 à 2%). Quant au sol, il est enrichi chaque année de 2 tonnes de matière organique par hectare (décomposeurs et humus).

Sur une surface de 1 hectare, soit 10000 m² on compte (données de l'équipe du professeur Duvigneaud) :
PRODUCTEURS :
Arbres et arbustes :
- 4 tonnes (t) de feuilles, fleurs et fruits
- 76 t de branches
- 180 t de troncs
- 54 t de racines
Plantes herbacées :
- 1 t de feuilles, fleurs et fruits
- 1 t de racines et organes souterrains
CONSOMMATEURS :
Grands mammifères : 2 kg
Petits mammifères : 5 kg
Oiseaux : 1,3 kg
Reptiles : 1,7 kg
Insectes : 1 kg
DÉCOMPOSEURS :
Faune du sol : 1 t (dont 600 kg de lombrics)
Flore du sol : 0,3 t
Humus : 1 à 15 t
Inventaire à la Prévert des forêts

Productivité primaire d'une forêt de nos contrées
La productivité primaire (bilan net) de notre forêt de feuillus tempérée est de l'ordre de 10 à 20 tonnes de matière sèche par hectare et par an.

Les écosystèmes forestiers contribuent à la formation d'un sol forestier 4 à 5 fois plus épais que celui d'une prairie, que les arbres protègent durablement en l'abritant de la pluie (un incendie entraîne ravinement et érosion). Une forêt agit par ses composantes stratigraphiques : les niches écologiques sont très diversifiées grâce à la répartition spatiale verticale des espèces de végétaux. La strate muscinale (mousses) ralentit le ruissellement et le lessivage du sol, tandis que les arbres pompent jusqu'à 10000 m³/ha/an. Ces usines photosynthétiques permettent, par leur formidable rejet d'oxygène moléculaire (6 à 20 t/ha/an), l'assainissement de l'atmosphère également favorisé parce qu'elles retiennent une grande partie des poussières (80 t/ha/an), et de certains composés toxiques contenus dans l'air. Enfin, une forêt, quand elle fait écran, réduit les bruits de 10 décibels par mètre d'épaisseur.

La limnologie est la science qui étudie les milieux aquatiques continentaux (eau douce, saumâtre ou salée) dont elle distingue deux classes :
- les milieux où l'eau s'écoule à plus ou moins grand débit, comme les ruisseaux, les torrents, les rivières et les fleuves, que l'on appelle milieux lotiques ;
- ceux où l'eau est plus ou moins stagnante, comme les mares, les étangs et les lacs, qu'on nomme milieux lentiques.
Parmi les écosystèmes les plus fragiles se trouvent les lacs et les étangs ; on comprend aisément qu'une pollution en rivière, où l'eau est constamment, donc rapidement, renouvelée, puisse paraître moins dramatique que si elle a lieu dans un milieu où l'eau circule trop lentement : lacs et étangs sont beaucoup plus vulnérables, en tout cas très sensibles aux moindres variations qui peuvent les affecter. Une bonne partie de nos milieux lentiques est malade et seul un bon diagnostic peut y remédier : la connaissance de l'écologie de tels milieux permet de trouver des solutions réparatrices qui, lorsqu'elles sont adaptées, laissent envisager une stabilisation durable de leur fonctionnement.
C'est la profondeur, plus que la superficie, qui permet la classification d'une étendue d'eau continentale en étang ou en lac : ce dernier prend son appellation dès lors que sa profondeur dépasse largement les 10 mètres (c'est un minimum), sinon c'est un étang (profondeur inférieure à 5 mètres en général). Un lac se compose de différentes zones ou régions biologiques dans lesquelles se répartit un grand nombre d'espèces aquatiques ou semi-aquatiques :
- la zone littorale de faible profondeur,
- la zone pélagique d'une profondeur supérieure à 2 m,
- l' épilimnion, soumis au vent, donc aux températures de l'air ambiant,
- le thermocline où la température chute de 1°C chaque fois qu'on s'enfonce d'un mètre,
- l' hypolimnion où l'eau n'est quasiment pas brassée, donc peu renouvelée ; que la lumière n'atteint plus à cause de la turbidité de l'eau (absence de producteurs) ; où la très faible teneur en oxygène moléculaire (O 2) limite la présence des êtres vivants à quelques espèces spécialisées (notamment détritivores et décomposeurs)

Dès lors que la profondeur excède les 10-15 m, s'installe un gradient thermique (ou stratification thermique) qui oppose l'épilimnion et l'hypolimnion ; cela n'existe pas dans les étangs dont la variation des températures de l'eau suit celle de l'air ; dans un lac profond, c'est plus complexe (voir schéma ci-dessous). Comme tous les écosystèmes, le milieu échange en permanence de l'énergie, de la matière et beaucoup de chaleur (milieu tampon) avec le milieu ambiant et les écosystèmes avoisinants. Comme nous le verrons, les étendues d'eau continentales sont de véritables pièges à énergie quand leur fonctionnement n'est pas affecté, ou quand l'homme fait intervenir le génie agricole afin de les rentabiliser (pisciculture) ou pour, simplement, les amender.


Le réseau trophique d'un lac (ou d'un étang) est extrêmement complexe mais, en le simplifiant bien, on retrouve le schéma classique d'un réseau trophique où les chaînes alimentaires débutent par les producteurs et se poursuit par les consommateurs de différents ordres ; sans oublier, même s'ils ne sont pas représentés ni dans la chaîne ni dans les pyramides écologiques, les décomposeurs qui permettent de boucler les différents cycles bio-géochimiques de la matière :
- les producteurs sont représentés par les végétaux chlorophylliens de la zone littorale et le phytoplancton. Grâce à leur capacité de capter l'énergie lumineuse du soleil - c'est le rôle de la chlorophylle de faire la photosynthèse -, les producteurs transforment le gaz carbonique (CO 2) qu'ils absorbent en sucres (glucides) et autres molécules organiques (lipides, protides...). Un producteur primaire a donc pour rôle essentiel de créer de la matière organique, riche en énergie calorique, à partir de matières minérales et de soleil ;
- les consommateurs sont classés par ordre croissant selon leur régime alimentaire. Les consommateurs de premier ordre (herbivores, phytophages, zooplancton) sont ceux qui ne se nourrissent que de producteurs ; les consommateurs de second ordre sont aussi des carnivores de premier ordre : ils se nourrissent de plancton (daphnies, poissons planctonophages), mais aussi d'insectes phytophages ; les consommateurs de troisième ordre (carnivores de deuxième ordre ou prédateurs, héron, brochet...) viennent clore cette chaîne alimentaire, se nourrissant des précédents. Bien qu'un lac ou un étang aient un excellent rendement énergétique, l'énergie captée en bout de chaîne devient insuffisante pour permettre l'installation d'un quatrième ordre de consommateurs (principe valable pour tous les modèles d'écosystèmes).

Comme dans tout cycle écologique élémentaire, les décomposeurs (détritivores compris, larves d'insectes, vers, champignons, bactéries...) jouent un rôle prépondérant dans le fonctionnement de l'écosystème lac ou étang : ils bouclent chaque cycle des éléments chimiques, notamment les cycles du carbone, de l'azote et du phosphore. Les différents groupes d'organismes qui recyclent à ce niveau la matière organique en la remettant sous sa forme minérale ne peuvent le faire correctement que sous certaines conditions d'oxygénation, c'est à dire là où l'oxygène moléculaire devient rapidement un facteur limitant en ce qui concerne l'entretien des phénomènes d'oxydo-réduction. La biomasse morte ainsi que les excréments rejetés par les animaux du biotope doivent subir une combustion oxydative cependant que sont libérés du gaz carbonique, des nitrates et des phosphates.
Les cycles reprennent (ou plutôt se poursuivent) lorsque ces derniers composés sont réutilisés dans les biosynthèses effectuées par les producteurs primaires (phytoplancton et végétaux chlorophylliens). Un défaut (manque d'oxygène par exemple) de décomposition peut entraver lourdement le fonctionnement écologique du milieu lentique, les nutriments de base (gaz carbonique, nitrates et phosphates) venant à manquer. L'alimentation est le facteur limitant par excellence dans de tels milieux ; il peut bloquer le déroulement des cycles dès le départ de la chaîne alimentaire en rendant impossible le transfert d'énergie (à la base, la lumière solaire) du phytoplancton aux divers prédateurs : la biomasse fait une chute dramatique au point que l'écosystème tout entier périclite de façon durable, et l'on pourrait craindre que l'équilibre ne soit à jamais rompu. C'est là que pourront intervenir les " médecins " du génie génétique (voir dernier chapitre de la première partie). Lorsqu'un étang piscicole est aussi bien géré qu'entretenu, il se comporte comme un formidable piège à énergie permettant une pêche annuelle allant jusqu'à 5 t/ha de poissons. Mis à part un agrosystème intensifié, peu d'écosystèmes terrestres peuvent se prévaloir d'une biomasse aussi importante.

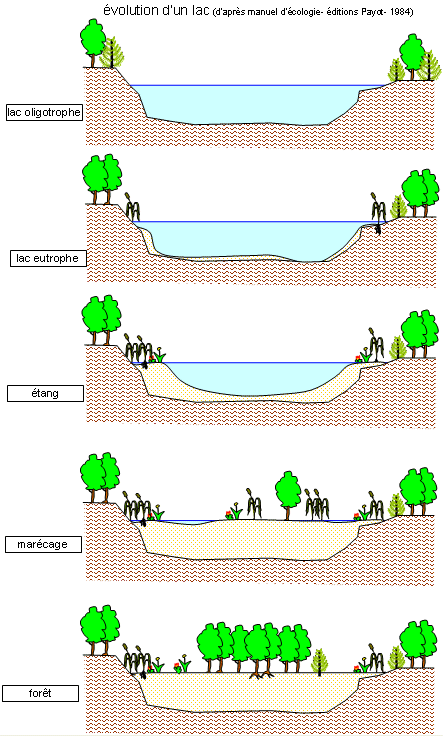
Évolution naturelle d'un plan d'eau
Un lac n'est pas une structure éternelle, il se transforme et évolue au rythme des différents cycles auxquels il est soumis. Un inévitable remplissage de terre (= atterrissement) finira par le combler intégralement ; tout dépendra bien entendu de sa superficie et de sa profondeur, initiales, de sa vitesse de sédimentation et de l'apport d'alluvions terrestres dans le temps. Pour bien comprendre, il faut imaginer la superposition de cycles multiples qui se déroulent tout au long de l'année et au cours de la vie du lac :
- cycles biologiques journaliers quand le plancton migre verticalement au gré des rythmes nycthéméraux (alternance jour/nuit),
- cycles thermiques saisonniers qui règlent le degré de stratification thermique (affirmée l'hiver et l'été, en phase d'homogénéisation de la température en automne et au printemps),
- cycles des phases évolutives propres au lac lui-même (formation et croissance, maturation, vieillissement, sénescence et disparition du lac).
Au départ, un jeune lac contient peu d'éléments nutritifs ; la croissance des producteurs - a fortiori celle des consommateurs, est donc limitée ; la biomasse qui s'en suit reste faible, la turbidité du lac est nulle (eau transparente) : ce sont les lacs oligotrophes de moyenne et haute montagne (lac Pavin, lac Chauvet), qui échappent jusque là aux pollutions diverses (agricole, industrielle et domestique).
Au fur et à mesure que la teneur en éléments nutritifs augmente, les producteurs (végétaux chlorophylliens et phytoplancton) croissent en volume et en quantité, l'apport de nutriments se faisant grâce aux bassins versants qui alimentent le lac en eau nouvelle chargée de minéraux. En mourant, la biomasse (ou nécromasse) produite se dépose au fond du lac (sédimentation) qui commence à se combler légèrement. Les décomposeurs y sont de plus en plus actifs, qui permettent aux cycles des éléments (oxygène, azote et potasse) de recommencer, inlassablement. La turbidité augmente de telle façon qu'on classe le lac parmi les lacs mésotrophes d'abord (lac Léman, lac du Bourget, lac d'annecy), eutrophes ensuite (lac de Grand-Lieu). Autant dire qu'il ne fait pas bon se baigner dans cette dernière catégorie !
Plus le lac se comble, moins la stratification thermique est affirmée ; elle tend à disparaître quand la profondeur de l'étendue d'eau est inférieure à 10 m. Ce phénomène entraîne une mauvaise oxygénation des sédiments, donc un resserrement de ce goulot d'étranglement du cycle de la matière, que représentent les décomposeurs, de moins en moins actifs. L'envasement du lac est tel qu'il prend l'appellation d'étang, puis de marécage... l'ancienne zone humide disparaît et fait place à un écosystème terrestre classique (lande... forêt pour achever la phase d'atterrissement). Bien sûr, la vie d'un lac se déroule sur des temps géologiques (plusieurs centaines de milliers d'années) indiscernables pour l'homme dont la vie est éphémère. On peut cependant lire l'histoire passée d'une étendue d'eau à ses différents stades en observant les marques fossilisées des anneaux concentriques d'associations végétales qui se resserrent au fur et à mesure que la profondeur diminue :
- Au delà de 5 m de profondeur se trouvent les hydrophytes (plantes strictement aquatiques comme les characées, les potamots, les nénuphars...) ;
- Sur la zone de balancement des eaux se trouvent les plantes semi-aquatiques, dont le feuillage est toujours hors de l'eau, mais qui prennent racines dans l'eau ; ce sont les hélophytes (joncs, phragmites -roseaux-, sagittaires, massettes...).
Chacune de ces zones possède également sa propre faune : oiseaux de la roselière, reptiles, amphibiens et petits mammifères de la zone littorale, insectes aquatiques de la zone pélagique (eau libre), canards divers et oiseaux limicoles... un nombre de niches écologiques incalculables qui résultent de la confrontation des écosystèmes aquatique et terrestre.
Eutrophisation d'un étang
Lorsqu'il est en équilibre dynamique (en pleine santé), un étang (un lac également) utilise de manière optimale l'énergie solaire lumineuse incidente ; sa biomasse est constante ; la matière organique y est recyclée de façon efficace et, pour finir, son fonctionnement est rendu stable grâce à certains phénomènes de régulation interne (photosynthèse, respiration, décomposition, reproduction...). La plupart du temps, l'homme gère et entretient les étangs pour éviter ou reculer leur comblement (atterrissement), en les rentabilisant par la pêche, mais aussi par un usage agricole, lorsque l'étang est vidangé tous les 3 à 5 ans.
Il arrive qu'un étang soit abandonné par l'homme pour une raison quelconque. Le simple fait de ne plus être ni vidé, ni curé, ni faucardé et pas plus fauché, entraîne des dérèglements dans ce très fragile écosystème. Une prolifération des producteurs primaires (herbes des prairies, plantes aquatiques et phytoplancton) s'en suit, augmentée par les apports minéraux provenant des terres environnantes, ce qui va accélérer le processus d'atterrissement : on parle d' eutrophisation de l'étang. Si rien n'est fait, quelques décennies " régleront son sort " à l'étendue d'eau, ce qui privera d'une aire d'étape les oiseaux migrateurs inféodés aux zones humides.
Plus l'étang perd en profondeur - par accumulation de la nécromasse -, plus la pellicule d'eau qui le constitue s'échauffe rapidement ; ce qui, aggravé encore par la turbidité de l'eau et la couleur sombre du plancher vaseux, empêche la lumière de pénétrer, limite donc la photosynthèse, et, par-là même, fait baisser la teneur des eaux en oxygène moléculaire. De nombreux poissons disparaissent peu à peu et leur décomposition, qui requiert aussi beaucoup d'oxygène, ne peut plus être effectuée par les bactéries et autres organismes aérobies (nécessitant de l'oxygène). La sédimentation se fait plus vite que la décomposition, car les micro-organismes anaérobies (contraire d'aérobie) qui prennent le relais ne sont pas de bons minéralisateurs ; de plus, leur métabolisme entraîne, dans le milieu, un dégagement de substances toxiques comme l'hydrogène sulfuré ou l'ammoniac. On parle alors de phénomène de dystrophysation en milieu lentique.
Au terme de la maladie, l'eau de l'étang n'est plus du tout oxygénée, la minéralisation de la matière organique est complètement bloquée, tel qu'elle s'accumule de plus en plus rapidement. Le milieu est devenu abiotique, c'est à dire impropre à la vie d'un milieu aquatique normal.

Retenons que la phase d' eutrophisation d'un étang connaît une prolifération de végétaux chlorophylliens qui, dans un premier temps, augmente la quantité d'oxygène dans l'eau. La biomasse zooplanctonique s'accroît tout naturellement, les poissons se multiplient... au point qu'on pourrait y voir un certain intérêt économique. Il s'agit d'une erreur car la nécromasse qui en découle devient si importante que son recyclage (re-minéralisation) par le benthos (êtres vivants du fond de l'étang) exige de plus en plus d'oxygène.
La phase suivante, ou dystrophysation, est caractérisée par un encombrement végétal tel que le soleil ne pénètre plus l'eau. La photosynthèse chute brusquement au point que la vie n'y est quasiment plus possible. Enfin, c'est l'entretien d'un étang, de ses rives et de ses berges, qui lui confère sa grande biodiversité : si l'on cesse de faucher les abords de l'étang, alors les espèces fragiles - car plus exigeantes -, comme les orchidées palustres, disparaissent au profit d'espèces plus ubiquistes comme les roseaux, les scirpes ou les massettes.
Les rivières font partie des milieux lotiques : leur eau est toujours courante. Celles de France, qu'elles trouvent leur source dans une région de basse altitude pourvue d'étangs, ou bien grâce aux torrents de montagne, sont toujours alimentées en eau. De fait, la plupart des espèces inféodées à ce milieu l'est strictement. Le réseau trophique, fort complexe, pousse ses extensions sur les berges (amphibiens, serpents, rongeurs...) ; il compte également des oiseaux prédateurs (busards, hérons, martin-pêcheurs...). Tout au long de l'année, le débit d'une rivière est variable ; il peut s'accélérer en fonction de la pluviosité, se mettre en crue ou en étiage. Cet écosystème est en constant réajustement et, même s'il est sensible à la moindre perturbation, il retrouve vite ses propres constantes paramétriques (température, teneurs en sels et en oxygène, pH...) : " l a teneur en oxygène de l'eau, la vitesse du courant et le débit jouent des rôles capitaux dans la répartition des communautés vivant le long du cours d'eau ".
Pour faciliter l'étude comportementale de la rivière, on la décompose en trois parties aux caractéristiques écologiques différentes :
♦ le crénon où se situe la source de la rivière : les paramètres biotiques et abiotiques restent assez constants tout au long de l'année, faune et flore sont relativement pauvres en ce qui concerne les rivières de montagne ;
♦ le rhitron est la partie supérieure de la rivière, aux eaux plus froides et plus rapides qu'en aval où le courant ralentit : la flore aquatique n'est pas abondante et la faune est dominée par les larves et les nymphes d'insectes (trichoptères, éphéméroptères) sensibles aux grosses variations de température ;
♦ le potamon se situe en aval de la rivière, c'est sa partie inférieure, aux eaux courant plus lentement ; de fait, la teneur en oxygène est moindre et, l'été, les températures augmentent en pouvant varier considérablement d'un jour à l'autre. On y trouve des espèces d'insectes peu sensibles aux variations du milieu (odonates, hémiptères) ainsi que des poissons peu exigeants, de la famille des cyprinidés essentiellement (carpes, tanches, brèmes, barbeaux...).
Comme dans tous réseaux alimentaires, le cycle de la vie d'une rivière débute avec des molécules minérales (nitrates, phosphates, potassium...) que végétaux et phytoplancton absorbent. Ces producteurs primaires sont ensuite consommés par le zooplancton, par des insectes phytophages et des poissons herbivores, eux-mêmes dévorés par des consommateurs secondaires. Consommateurs primaires et secondaires servent encore de nourriture aux consommateurs d'ordres supérieur (brochet, busards, hérons...). Le cycle se poursuivra une fois le travail des détritivores (larves de chinoromides, vers divers...) et des décomposeurs (bactéries, champignons) effectué. Ces derniers terminent le processus de transformation des matières organiques (cadavres d'animaux et de plantes, excréments) en matières minérales (nitrates, phosphates, potassium)...
Quantité d'échanges a lieu entre le potamon et le rhitron : les insectes remontent en amont pour pondre leurs œufs alors qu'une partie de leurs larves sera emportée par le courant vers l'aval ; de même, la rivière interfère avec les milieux terrestre et aérien, ce qui rend cet écosystème à la fois particulièrement dynamique et atypique. Le " continuum fluvial " se traduit par des liens physiques entre amont et aval, grâce au travail hydraulique, érosif et sédimentaire des eaux de gravité, mais aussi par des relations alimentaires et comportementales (va et vient des poissons pour frayer) ; de même, la rivière reste liée, au moment des crues plus ou moins importantes, à l'ensemble de la plaine alluviale avec laquelle s'effectuent des échanges bénéfiques mutuels. Enfin, cette dynamique se poursuit par la solidarité verticale qui lie le fond de la rivière et ses eaux de surface (brassage et oxygénation des différentes couches d'eau grâce aux remous occasionnés par les pierres).
Ce cycle d'une grande complexité est entretenu par la pluviosité et la fonte des glaciers ou des neiges sommitales. Si les précipitations venaient, à cause du changement climatique tant annoncé, à se raréfier, on pourrait envisager un arrêt saisonnier total (été) de certains cours d'eau de nos régions (pas forcément qu'au Sud). Le phénomène pourrait alors, s'il atteignait un paroxysme avec 6 ou 7 °C au dessus des moyennes actuelles thermiques - comme du temps des dinosaures par exemple -, toucher même les grands fleuves au plus fort de l'été. Cela reste purement spéculatif...

La plupart des tourbières de France (20000 ha de superficie totale) se trouve dans les régions montagneuses (1000 m d'altitude en moyenne) ; elles sont des reliquats du dernier âge glaciaire qu'elles trahissent, le fameux épisode Würm qui donnait de son maximum il y a 20000 ans. Chaque dépression suffisamment importante et remplie d'eau pouvait donner naissance à une tourbière, du moment que la décomposition des matières mortes (végétales et animales) restait bloquée par la froideur du climat et tant que l'oxygénation du milieu restait insignifiante ; c'est pourquoi l'eau, le plus souvent permanente, devait y stagner par manque de renouvellement. La faune et la flore, très particulières, sont relativement proches de celles que l'on rencontre dans la toundra arctique.

La biodiversité d'une tourbière est remarquable tant cette dernière s'apparente à un îlot de vie boréale. Certaines espèces illustrent bien les phénomènes d'adaptation à ces milieux extrêmes :
◊ aux premiers stades de la tourbière, des plantes carnivores (drosera, utriculaire) consomment des insectes pour compenser un déficit en azote dans le sol ;
◊ dans les tourbières bombées, les laîches et la molinie forment des touradons (sorte de mottes de hauteur variable) pour exposer leurs parties végétatives à la pleine lumière ;
◊ les sphaignes, qui sont les végétaux représentatifs de la tourbière, ont d'énormes capacités d'absorption d'eau et de croissance; ce sont elles qui forment le " sol " en se fossilisant ; proches des sphaignes sont les mousses qui les accompagnent (lycopode, polytriche, hypnum) ;
◊ certaines larves de libellules vivent 3 ans dans ces milieux pauvres avant leur métamorphose ;
◊ des végétaux et des insectes " nordiques " persistent grâce aux tourbières de montagne sous nos latitudes : myrtille des lapons, andromède à feuilles de polium, lycène helle (papillon), libellule dubia

Au fil du temps - un temps géologique -, les végétaux meurent mais ne sont pas décomposés ; ils s'accumulent en couches successives, dans une eau extrêmement pauvre en sels minéraux et dont le pH avoisine 4 (très forte acidité). En profondeur à lieu le phénomène de tourbification, c'est à dire la métamorphose des restes organiques en tourbe*, une roche carbonée (combustible) contenant environ 50% de son poids sec en carbone. Lorsque la profondeur le permet avec le comblement inéluctable de la tourbière, les sphaignes gagnent peu à peu le centre de l'étendue d'eau qui se réduit à une " peau de chagrin "...

...et finissent par l'envahir toute entière pour achever son évolution par un atterrissement total de la tourbière. Lorsque celle-ci a atteint son dernier stade, les arbres peuvent la coloniser et la peupler complètement. Parfois, la trace de sa présence est complètement effacée en surface ; on ne peut en retrouver les vestiges qu'en faisant des prélèvements profonds, par carottage. C'est ainsi que les palynologues, en étudiant les grains de pollens qui se sont progressivement déposés à sa surface, fossilisés également, sont capables de retracer l'histoire de la tourbière, relativement aux climats qui se sont succédés dans le temps.

* tourbe : " un sédiment, un sol ; il y en a différents types (variations de composition et de couleur, de texture) ; elle abrite des vestiges de temps lointains (bois, élytres de coléoptères...) visibles dans les carottes ; les grains de pollen et autres restes végétaux permettent de reconstituer les climats et végétations passés. Les " archives naturelles " des tourbières nous ouvrent les portes du passé, souvent depuis la fin de la dernière glaciation (12000 ans) ! La tourbe a aussi été utilisée pour différents usages : combustible, supports de culture, pansements... entraînant le développement de métiers, de coutumes, d'outillages spécifiques. Enfin, la tourbe stocke efficacement et durablement le carbone. "
